Unit 3. Life: From Building Blocks to Biomes
What are the building
blocks of life?
Even though there are more than
92 natural elements found on Earth, almost all life forms on Earth are primarily
made up of only four basic elements: hydrogen, oxygen, nitrogen, and carbon. An
element is a substance that is made
of only one type of atom. When two or more atoms join together through chemical
or ionic bonding, they make up a new molecule.
Elements can combine together in many different ways to form thousands of
different compounds. Compounds are generally thought of as larger
aggregates of molecules. Most elements combine to form compounds through
covalent bonds, with carbon containing four bonds, oxygen two bonds and
hydrogen one bond. Water for example is a simple molecule composed of two
hydrogen atoms, and one oxygen atom bonded together; however, water makes up a
significant proportion of living organisms on Earth, and it is important because
no organism can be biologically active without it.
Nitrogen is also essential for
life because nitrogen-based compounds make up DNA or deoxyribonucleic acid,
which is the blueprint for all life on Earth. DNA facilitates building all of
the other larger structures necessary for life to function and replicate.
Nucleotides are the basic chemical unit in a nucleic acid, which are the
building blocks of DNA and RNA (ribonucleic acid). Nucleic acids are formed
from nucleotides, which are made of a pentose sugar, a phosphate group and a
nitrogen-containing base. Only four nitrogenous bases are found in DNA:
adenine, cytosine, guanine, and thymine. Adenine (A) and guanine (G) are larger
bases called purines and thymine (T) and cytosine (C)
are smaller bases called pyrimidines. A purine such as adenine must always pair with a pyrimidine (thymine) in the DNA strand. This complimentary
pairing creates the distinctive double helix structure of DNA (Figure 3.1) and
makes possible the replication of DNA.
Life is made of organic
compounds, which contain the element carbon. Carbon plays a key role in life,
because it links together other elements to form large and complex molecules.
In the DNA double helix, the nucleotide sequence codes for the building of
amino acids. Amino acids are a group of twenty carbon-based compounds that have
a carboxyl group and an amino group (nitrogen containing) bonded to the same
carbon atom. They are important for life because they serve as the building
blocks of protein molecules that make up living organisms. Each type of amino
acid has a unique side chain that gives it distinct chemical properties, which
adds to the diversity of structures that can be made from amino acids. Amino
acids can be linked together through a condensation reaction to form
polypeptide chains. One or more polypeptides chains can then form a protein.
The number of polypeptides and the structure of the proteins are determined by
the sequence of nucleotides in the DNA molecule.
Proteins are extremely diverse
and essential for life as they are the building blocks for cell construction,
maintenance, and reproduction in living organisms. Structural components of
life such as cell membranes, muscles, and tissues are all made from proteins.
In addition, enzymes, which regulate chemical reactions in all living systems,
are made of proteins. Carbohydrates and fats are two other key components of
living organisms that are necessary for life. Carbohydrates and fats primarily
serve to supply energy for construction, maintenance, and reproduction costs in
living organisms. In addition, fats are important as a component of cell
membranes.
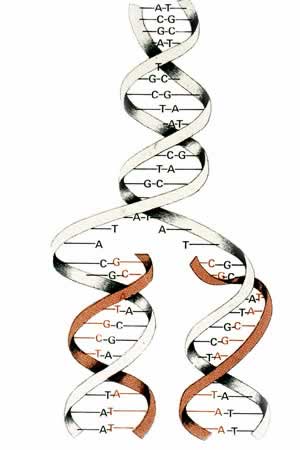
Figure 3.1
Structure of DNA
http://www.genelex.com/paternitytesting/images/dna-molecule.jpg
What are genes and what
do they do?
Genes are made up of DNA and
are found on chromosomes. Genes are considered to be the primary unit of
heredity. Genes determine the different traits or characteristics that an
organism displays, and alternative forms of a gene are called alleles. For example,
if you inherited your mother’s curly hair you may have received her dominant
allele for the gene that is expressing that trait.
To better understand the
terminology associated with genes, we will use classic Mendelian
genetics. Mendelian genetics is the standard pattern
of inheritance for sexually reproducing organisms and was discovered by Gregor Mendel, an Austrian monk that used the garden pea to
study patterns of inheritance. He determined that physical expression of traits
(or the phenotype) was determined by the genetic make up (or the genotype) of
the individual.
Mendel also determined that
pairs of alleles were responsible for an organism’s genotypic make up, and the phenotypic
expression of the traits associated with that genotype. For example, Mendel
found that if two alleles were identical, the individual was homozygous for
that gene. Let’s examine the peppered moth (Biston
betularia) as a real world example of how genes
might work in nature (Table 3.1). In the case of the peppered moth, the allele
for light body color has a stronger phenotypic expression than that of the dark
body color; therefore, the light color is considered to be dominant and the
dark color is recessive. If two alleles are different, the individual is
heterozygous, or has two different alleles for a particular gene. When moths
are heterozygous, the dominant allele (in this case the light body color) has
such a strong phenotypic expression that it conceals the presence of the weaker
or recessive allele (the dark body color).
If we cross a dark bodied male
(dd recessive) with a light bodied female (LL
dominant) moth we will end up with all light colored moths in the F1
or first generation of offspring (Table 3.1). This is because all of the
offspring will be the heterozygous (Ld) genotype.
Next, if we cross the two heterozygous moths (Ld x Ld) from the F1
generation, we will end up with three light bodied moths (LL, Ld, Ld) and one
dark bodied moth (dd) in the F2 or second
generation. This is because half of the moths will be homozygous, one dominant
and one recessive (LL and dd) and half will be
heterozygous (Ld) like the F1 generation.
|
Moth Hybridization
|
Generation
|
# of Light Phenotype
(Genotype)
|
# of Dark Phenotype
(Genotype)
|
|
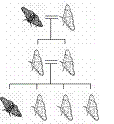
|
F0 (parents)
|
1 Light
(LL)
|
1 Dark
(dd)
|
|
F1
|
All Light
(Ld)
|
0 Dark
(Ld)
|
|
F2
|
3 Light
(1 LL, 2 Ld)
|
1 Dark
(1 dd)
|
Table 3.1
Phenotypic expression of genotypes in the peppered moth
http://www.wwnorton.com/college/anthro/bioanth/ch2/
Evolution is dependent on
changes in the genetic frequencies in a population, not in the individual.
Allele frequencies are important for studying genetic variation in the gene
pool, or the total of genetic information present in a population. Genetic
variation then, is central to understanding evolutionary change in populations.
We can determine mathematically whether a population is changing, or whether it
is at equilibrium using the Hardy-Weinberg equation. The Hardy-Weinberg rule
tells us that if a population is at equilibrium it will have the same genotypic
frequencies from generation to generation, and that allele frequencies will
remain the same unless some selection pressure acts to change them. There are
many types of selection pressures that may change the genotypic frequencies in
a population. Some of these selection pressures include: genetic mutations,
genetic drift as a result of bottlenecks or the founder effect, migration, and
lastly natural selection.
What is natural
selection and how does it work?
Natural selection is the
differential selection of genotypes and is part of evolutionary theory, which
was developed by Charles Darwin. Evolution can be considered as change in the genetic
composition of a population with the passage of each generation. One of the
mechanisms of evolutionary change that Darwin
developed was natural selection.
There are three basic types of
natural selection (Figure 3.2). Under diversifying (disruptive) selection, both
extreme phenotypes are favored at the expense of intermediate varieties. This
is uncommon in nature, but it suggests a mechanism for species formation
without geographic isolation. The second type of natural selection is
directional selection. Under directional selection, individuals at one end of
the phenotypic distribution survive or reproduce better than those with the
other phenotype. Thus, the frequency distribution of the trait in the evolved
population (or the second generation) is shifted toward the more favorable
trait. This is what we typically think of as natural selection. The third type
of natural selection is stabilizing selection. Under stabilizing selection,
extreme varieties from both ends of the frequency distribution are eliminated.
This is one of the most common forms of natural selection. Human birth weight
is a good example of this type of selection, with unusually large babies and
unusually small babies being selected against and medium-sized babies being
selected for in the population.
Figure 3.2
Types of natural selection
For natural selection to occur,
two requirements must be met: First, traits must be heritable. For example,
body color in moths or beak size in birds, which is what Darwin studied, are inherited traits. Second,
possession of a trait must then produce differential survival or reproduction
rates in the population of organisms. Unless both these requirements are met,
adaptation by natural selection cannot occur. Let’s examine the peppered moth
as an example of directional natural selection.
Prior to the 1800’s in England, the typical peppered moth
was light bodied with black spots. Dark colored moths were rare and were
therefore collectors' items, with the first one being found in 1819. During the
Industrial Revolution, soot and other industrial pollution darkened tree trunks
and killed off lichen communities that lived on the trees. This made the light
colored moths more visible to bird predators and caused the light-colored moths
to become differentially selected by bird predators. The selective predation by
birds, which favored camouflage coloration in the moth and was in turn
dependent on the color of the tree-trunks, caused the light-bodied moths to
become more rare and the dark-bodied moths to become
more abundant. By 1886 dark-bodied moths were far more common than
light-colored moths, illustrating rapid evolutionary change in the population.
Eventually light moths were common in only a few locations, far removed from
industrial areas.
How
does speciation happen?
Historically there have been two
main ways to differentiate between species. One way is the morphological
species concept, which differentiates between species based on anatomical
morphology, or the shape and look of the body and its parts. Another way to
differentiate between species is based on the biological species concept, or
the ability to interbreed. This methodology basically assumes that if organisms
can interbreed and have biologically viable offspring they are the same
species, and if they cannot produce viable offspring they are different
species.
Distinct species appear when a
single lineage splits into two populations that are no longer capable of
inter-breeding. Usually this occurs as a
result of geographic isolation, due perhaps to mountain building, climate
change, or when a river cuts a deep chasm that prevents movement of individuals
between the isolated populations. This is called allopatric
speciation or speciation due to geographic isolation, and is the most common
form of speciation (Figure 3.3). Genetic differences accumulate by chance and
may be reinforced by different selection pressures in the two locations. Given sufficient time, the two populations
will accumulate enough differences that they are unable to produce viable
offspring if they come back into contact.
Reproductive isolation is the key attribute of species-hood, and it can
arise from prezygotic (those that prevent conception)
or postzygotic (those that prevent the normal
development and birth of a healthy individual) isolation mechanisms. Prezygotic isolation mechanisms prevent the breeding
process from occurring between two populations in the first place. For example,
if the two new species have different breeding seasons, say early and late
spring, they may never be ready to breed at the same time and so experience a
temporal prezygotic isolation mechanism. On the other
hand, if two populations were not separated by their breeding period, they may
occupy different habitats or have different courtship displays. Distinctive courtship behaviors allow
individuals to ensure that they breed with other individuals of the same
species, and thus avoid wasting a breeding opportunity.
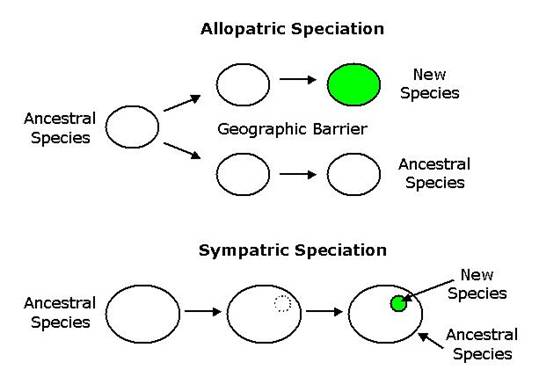
Figure 3.3
Allopatric and Sympatric speciation
Postzygotic
isolation mechanisms include the failure of sperm and egg to unite and begin
development, the inability of hybrid progeny to develop to the point of birth,
or the birth of individuals that are less able to feed, grow and compete than
non-hybrids. One form of postzygotic isolation is
hybrid inviability where the zygote develops
abnormally and is aborted. Hybrid sterility is also a form of postzygotic isolation where the zygote may develop and be
healthy; however, the individual born is sterile because the different
chromosomes inherited from its parents do not pair and cross over properly
during meiosis, thus the individual cannot produce viable offspring of their
own. Another postzygotic isolation mechanism manifests
as reduced reproductive fitness, which may occur directly with the hybrid
individual or may occur over a few generations, eventually leading to inviable offspring in the hybrid lineage. Thus there are several ways in which
reproductive isolation is achieved, and these speciation processes can occur
rapidly in some organisms (like bacteria) or over hundreds or thousands of
years in other organisms.
What is a terrestrial
biome and why does the Earth have them?
When we combine all of the trophic levels and the web of their interactions with the abiotic environment, we refer to that as an ecosystem, and ecosystems are combined
into biomes. Biomes are Earth’s major
terrestrial ecosystems, or large biotic regions of Earth influenced by
precipitation, temperature, humidity, latitude, and topography. Biomes are
often grouped by the characteristics of the major vegetation types, animals,
and climate that is associated with the biome location. The classification of
terrestrial biomes is based upon the structure (or the physiognomy) of the
vegetation, and the names generally give us information about the dominant
vegetation type in the biome, as well as the type of climate that created and
maintains the biome. For example, cold deserts, hot deserts, temperate
rainforests, and tropical evergreen forests are all examples of biomes that
give us information about the climate and dominant vegetation type associated
with the biome. Note that the major
units of aquatic ecosystems are not characterized by the term “biome”, mainly
because the fundamental habitat is all water, whether it is salty or
fresh. However, just as for the
terrestrial ecosystems, aquatic systems are affected strongly by climate and
their location on Earth.
The spatial structure of
terrestrial biomes corresponds to the Earth’s major climatic zones (Figure
3.4). The Earth’s climate is central in determining the annual temperature
ranges, the amount of precipitation and the potential evapotranspiration
in different locations on Earth. Because soil is formed from the geologic
parent material and from physical and chemical weathering (which is dependent
on temperature and rates of precipitation) in a particular location, biomes are
also found to be related to the soil types found on Earth.
Biomes are generally found
parallel to the lines of latitude on Earth, which also indicates that biome
distribution is related to climate (Figure 3.5). One exception to this occurs
in mountainous regions. Mountain ranges differ in their pattern of temperature and
precipitation compared to lowland areas at the same latitude. As elevation
increases, temperature decreases and precipitation is generally more readily
available. Therefore, ecosystems in mountainous areas will be more similar to
biomes found in polar-regions. This change in biome type will become more
extreme as elevation increases. For example, Hopkin’s
bioclimatic law tells us that if you are in the northern hemisphere and
increase your elevation by just 1000 feet, you have essentially moved the
ecosystem equivalent of 100 miles north in latitude (Figure 3.6).
Biome types can be broken down
into a large number of specific categories. However, some of Earth’s classic
biomes include tundra, taiga (coniferous forests), tropical rainforests,
deciduous forests, grasslands, and deserts. Tundra occurs in the Arctic and in high mountainous regions at all latitudes.
Tundra is characterized by subsoil that is permanently frozen, with high
precipitation rates and low temperatures. Temperatures in tundra biomes are too
cold for trees; therefore, the dominant vegetation types are mosses, lichens,
grasses and low growing perennial herbs. The taiga or boreal forest biome is
located south of the Arctic tundra and at lower elevations than the alpine
tundra ecosystem. Boreal forests are characterized by long winters with heavy
snow and short warm summers, and they tend to have low species diversity with
coniferous evergreen trees as the dominant vegetation type. The deciduous
forest biome is located south of the taiga on the eastern side of North America
and Asia, and in Western Europe. Deciduous
forests are dominated by broadleaf trees and have temperate climates with ample
rainfall. Tropical Rain Forests generally are found in equatorial regions with
abundant rainfall and warm temperatures. Tropical rainforests have the highest
diversity of plant and animal species on the planet. Grasslands are found in
many regions on Earth where rainfall is limited and unevenly dispersed
throughout the year. Grasslands are also species rich. Many agricultural
regions of the world were once grasslands. Deserts are found in areas with
extreme temperature fluctuations and extremely low rainfall.
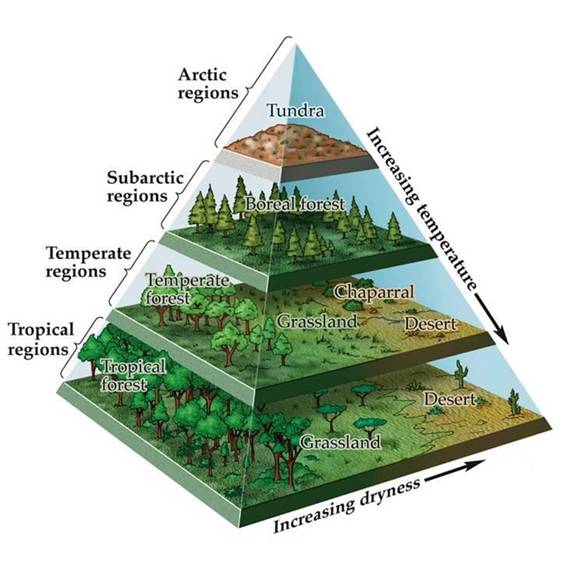
Figure 3.4
Temperature and precipitation related to biomes
http://www.csus.edu/indiv/l/loom/ecol/terrest%20biomes.JPG
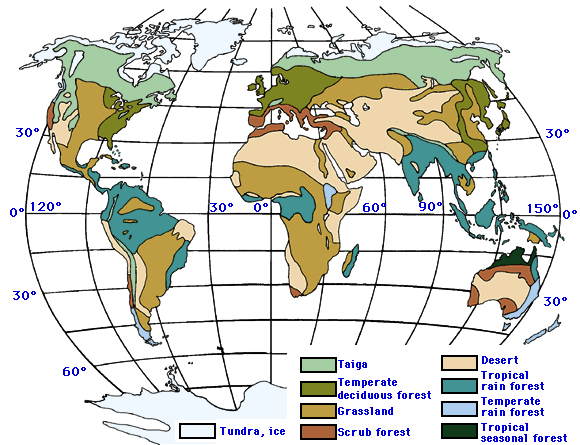
Figure 3.5
Biomes and latitude
http://www.agen.ufl.edu/~chyn/age2062/lect/lect_28/40_31A.GIF
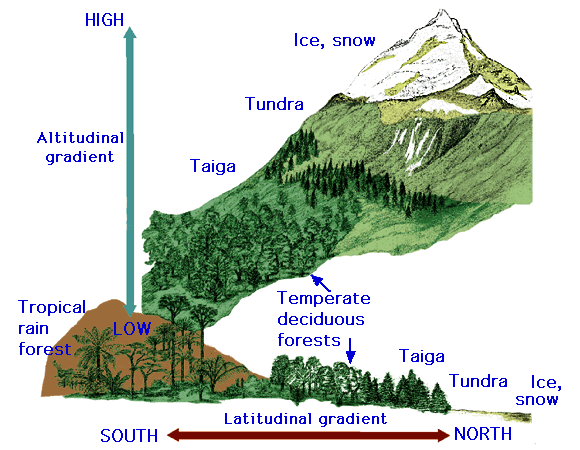
Figure 3.6
Biomes and altitude
http://www.agen.ufl.edu/~chyn/age2062/lect/lect_28/40_38.GIF
What effect does human consumption
have on the life system?
Currently, the human global
population is greater that 6.6 billion people and it is
projected to reach 9 billion by 2050. Because the human population has been
increasing at such a rapid rate for the past one hundred years or more, we are
using greater quantities of resources to sustain our needs. Worldwide there
exist fewer than 1.8 biologically productive hectares per person. According to
NASA, humans alone now consume approximately 20% of all the net primary production
(NPP) that the Earth produces every year (Figure 3.7).
Ecological footprint analysis
calculates the amount of ecologically productive land area that it takes to
sustain a population by accounting for the food, energy, and water used by the
population, as well as the waste generated in the process. To be ecologically
sustainable, each person living on the planet should require less than 1.8
hectares of land for their total ecological footprint. However, the average
American’s ecological footprint is roughly eight times that amount! Human
consumption rates are influenced by the number of people in a population, the
per capita consumption rate, and the technology that is available, which varies
greatly by country and region.
Globally, humans are consuming
renewable resources at a faster rate than the Earth’s ecosystems can replenish
them. Due to this over-consumption, approximately 60% of world’s ecosystems
have been degraded. The 2005 Footprint of Nations report determined that the
current global population is consuming 39% more of Earth’s biological capacity
than is ecologically sustainable. This unsustainable rate of consumption has
negative consequences for many of the plant and animal species with which
humans coexist. For example, human consumption pressures have resulted in
irrevocable losses in Earth’s biodiversity with approximately 10-30% of all amphibian, bird, and mammal species threatened with
extinction. Furthermore, the current rate of species loss is 100 to 1,000 times
greater than the natural extinction rate. In addition to directly consuming
resources, humans affect biological diversity by disrupting and destroying
habitats, introducing exotic species and pests, as well as contaminating
ecosystems with waste and pollution.
Much of the ecosystem
degradation and species loss on Earth is associated with high levels of
deforestation to keep up with increasing population growth rates and
consumption demands. Logging for timber, paper, and fuel, as well as land
conversion for agriculture are the primary causes of deforestation. Forest ecosystems house nearly 70% of the world’s
terrestrial biodiversity. Unfortunately, the World Resources Institute
estimates that more than 80% of the Earth’s natural forests already have been
damaged or destroyed with 12 million hectares of forest being cleared annually.
In addition, the world’s tropical forests are highly endangered and at the
current deforestation rate some impoverished African, Asian, and Central
American countries are expected to completely destroy their remaining tropical
forests in the next 5-10 years.
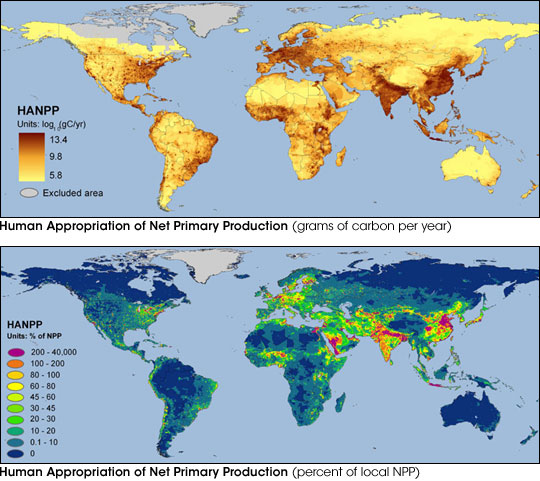
Figure 3.7
Human appropriation of global NPP
http://earthobservatory.nasa.gov/Newsroom/NewImages/images.php3?img_id=16586
Last updated: 10/23/2006 6:40 PM
